1.1 Cell Theory, Basic Properties of Cells & Origin of Eukaryotic Cells
Types of Microorganisms
Life takes many forms, from giant redwood trees towering hundreds of feet in the air to the tiniest known microbes, which measure only a few billionths of a meter. Humans have long pondered life’s origins and debated the defining characteristics of life, but our understanding of these concepts has changed radically since the invention of the microscope. In the 17th century, observations of microscopic life led to the development of the cell theory: the idea that the fundamental unit of life is the cell, that all organisms contain at least one cell, and that cells only come from other cells.
Despite sharing certain characteristics, cells may vary significantly. The two main types of cells are prokaryotic (lacking a nucleus) and eukaryotic (containing a well-organized, membrane-bound nucleus). Each type of cell exhibits remarkable variety in structure, function, and metabolic activity (Figure 1). Unit 1 will focus on the historical discoveries that have shaped our current understanding of cells, including their origins.

| Unit 1, Topic 1—To Do List | Suggested Average Time to Complete (min) |
|---|---|
| Read, make summary notes, and complete the self-check questions for Foundations of Modern Cell Theory. | 20 |
| Complete Learning Activity: Cell Theory. | 17 |
| Complete Learning Activity: How Science Builds. | 20 |
| Read, make summary notes, and complete the self-check questions for Basic Properties of Cells. | 20 |
| Complete Learning Activities: Parts of a Cell; Cell Comparisons; Cell Structure and Function. | 30 |
| Read, make summary notes, and complete the self-check questions for Origin of Eukaryotic Cells. Pay particular attention to the Case Study “What does “slug power” have to do with the origins of Chloroplasts?” | 20 |
| Complete Learning Activity: Endosymbiotic Theory. | 10 |
| ✮ Complete Learning Activity: Endosymbiotic Theory Online Interactive. Note that this activity is included as an option in the Unit 1 Assignment. | 30 |
Foundations of Modern Cell Theory
Section Learning Objectives
By the end of Unit 1, Topic 1, you will be able to:
- Explain what made the discovery of cells possible.
- Explain the key points of cell theory and the individual contributions of Hooke, Schleiden, Schwann, Remak, and Virchow.
- Explain how science builds.
Discovery of Cells
In this section, you will read about experiments that revealed secrets of cell and molecular biology, many of which earned their researchers Nobel and other prizes. But let’s begin here with a Tale of Roberts, two among many giants of science in the Renaissance and Age of Enlightenment, whose seminal studies came too early to win a Nobel Prize.
The English scientist Robert Hooke first used the term “cells” in 1665 to describe the small chambers within cork that he observed under a microscope of his own design (Figure 2). Hooke was one of the earliest scientists to study living things under a microscope (investigate Hooke’s drawings and microscope). The microscopes of Hooke’s day were not very strong and magnified images by 30x. However, Hooke was still able to make an important discovery. When he looked at a thin slice of cork under his microscope, he was surprised to see what looked like a honeycomb. To Hooke, thin sections of cork resembled “Honey-comb,” or “small boxes or bladders of air.” He noted that each “Cavern, Bubble, or Cell” was distinct from the others (Figure 2).
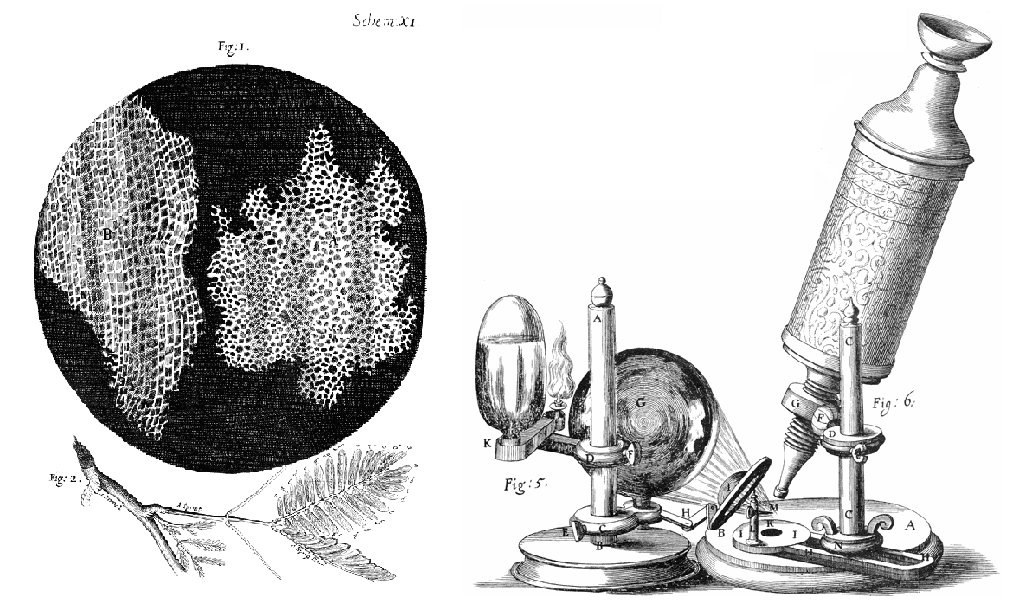
Soon after Robert Hooke discovered cells in cork, a Dutch tailor, Antonie van Leeuwenhoek (1632-1723), made other important discoveries after improving his own microscope. As a tailor, van Leeuwenhoek used lenses to examine cloth, and it was probably this incentive that led to his interest in lens making. His microscope was more powerful than other microscopes of his day (magnifying images by 270x), and it was almost as strong as modern light microscopes, even though it had a single lens. Using his microscope, Leeuwenhoek discovered tiny living animals, such as rotifers. Leeuwenhoek also discovered human blood cells. He even scraped plaque from his own teeth and observed it under the microscope!
Self-Check
- What type of material is cork? Do you know where cork comes from?
Show/Hide answer.
It’s from tree bark. At the time, Hooke was unaware that the cork cells were long dead and therefore, lacked the internal structures found within living cells.
- What do you think Leeuwenhoek saw in the plaque? He saw tiny living things with a single cell that he named animalcules (“tiny animals”).
Show/Hide answer.
Bacteria. Today, we call Leeuwenhoek’s animalcules bacteria.
Source: 2.2 Foundations of Modern Cell Theory (Bruslind/OpenStax)
Cell Theory
As described above, the discovery of the cell was made possible by the invention of the microscope. The microscopes we use today are far more complex than those used in the 1600s by Antonie van Leeuwenhoek. Later advances in lenses, microscope construction, and staining techniques enabled other scientists to see some components inside cells. Despite Hooke’s and van Leeuwenhoek’s early description of cells, their significance as the fundamental unit of life was not yet recognised until nearly 200 years later.
At this time, the German botanist Matthias Jakob Schleiden (1804–1881) was the first to recognise that all plants, and all the different parts of plants, are composed of cells. Visualizing plant cells was relatively easy because plant cells are clearly separated by their thick cell walls. Schleiden believed that cells formed through crystallization, rather than cell division. While having dinner with zoologist Theodor Schwann (1810–1882), Schleiden mentioned his idea. Schwann, who came to similar conclusions while studying animal tissues, quickly saw the implications of their work. In 1839, he published “Microscopic Investigations on the Accordance in the Structure and Growth of Plants and Animals,” which included the first and second statements of the cell theory: “All living things are made up of one or more cells,” and “the cell is the structural unit of life.”
In the 1850s, two Polish scientists living in Germany pushed this idea further, culminating in what we recognise today as the modern cell theory. In 1852, Robert Remak (1815–1865), a prominent neurologist and embryologist, published convincing evidence that cells are derived from other cells as a result of cell division. Based on this realization, Virchow extended the work of Schleiden and Schwann by proposing that living cells arise only from other living cells. However, this idea was questioned by many in the scientific community because most people, scientists included, believed that nonliving matter could spontaneously generate living tissue. Three years later, Rudolf Virchow (1821–1902), a well- respected pathologist, published an editorial essay entitled “Cellular Pathology,” which popularized the concept of cell theory using the Latin phrase omnis cellula a cellula (“all cells arise from cells”), which is essentially the third tenet of modern cell theory (3). Given the similarity of Virchow’s work to Remak’s, there is some controversy about which scientist should receive credit for articulating cell theory. See the following Science and Plagiarism feature below for more about this controversy.
The expanded version of the cell theory can also include the following (4):
- Cells carry genetic material passed to daughter cells during cellular division.
- All cells are essentially the same in chemical composition.
- Energy flow (metabolism and biochemistry) occurs within cells.
Eye on Ethics: Science and Plagiarism
Rudolf Virchow, a prominent, Polish-born, German scientist, is often remembered as the “Father of Pathology.” Well known for innovative approaches, he was one of the first to determine the causes of various diseases by examining their effects on tissues and organs. He was also among the first to use animals in his research and, as a result of his work, he was the first to name numerous diseases and created many other medical terms. Over the course of his career, he published more than 2,000 papers and headed various important medical facilities, including the Charité – Universitätsmedizin Berlin, a prominent Berlin hospital and medical school. But he is, perhaps, best remembered for his 1855 editorial essay titled “Cellular Pathology,” published in Archiv für Pathologische Anatomie und Physiologie, a journal that Virchow himself cofounded and still exists today. Despite his significant scientific legacy, there is some controversy regarding this essay, in which Virchow proposed the central tenet of modern cell theory—that all cells arise from other cells. Robert Remak, a former colleague who worked in the same laboratory as Virchow at the University of Berlin, had published the same idea three years before. Though it appears Virchow was familiar with Remak’s work, he neglected to credit Remak’s ideas in his essay. When Remak wrote a letter to Virchow pointing out similarities between Virchow’s ideas and his own, Virchow was dismissive. In 1858, in the preface to one of his books, Virchow wrote that his 1855 publication was just an editorial piece, not a scientific paper, and thus there was no need to cite Remak’s work. By today’s standards, Virchow’s editorial piece would certainly be considered an act of plagiarism, since he presented Remak’s ideas as his own. However, in the 19th century, standards for academic integrity were much less clear. Virchow’s strong reputation, coupled with the fact that Remak was a Jew in a somewhat antiSemitic political climate, shielded him from any significant repercussions. Today, the process of peer review and the ease of access to the scientific literature help discourage plagiarism. Although scientists are still motivated to publish original ideas that advance scientific knowledge, those who would consider plagiarizing are well aware of the serious consequences. In academia, plagiarism represents the theft of both individual thought and research—an offense that can destroy reputations and end careers.
Despite that we are told repeatedly in school to conduct ourselves with academic integrity, professional researchers do not always behave this way. Retraction Watch is a blog about scientific publications that had been suspected of a lack of integrity, such as fabricated data, and have been retracted. The lack of integrity not only damages the reputation of the scientists who had their papers retracted but also harms other researchers. It may take years for a retraction to be decided and publicized. By the time a paper is retracted, other researchers may already have spent large amounts of money and time in pursuing a topic that had been falsified. Worse yet are the fraudulent papers that have not yet been identified because people may unknowingly use the false information in subsequent papers and hence propagate information that is actually wrong. Therefore, it is important for everyone to produce work with integrity, to protect oneself and others.
The panels below (Figure 3) show (a) Rudolf Virchow (1821–1902) popularized the cell theory in an 1855 essay entitled “Cellular Pathology.” and (b) Robert Remak (1815–1865) who first published the idea that all cells originate from other cells in 1852 and who was a contemporary and former colleague of Virchow (Kisch 1954; Harris 2000; Webster 1981; Zuchora-Walske 2015).

Learning Activity: Cell Theory
- Visit the following sites to learn more about van Leeuwenhoek’s contributions to the Cell Theory. The videos will provide you with a visualization of van Leeuwenhoek’s important contributions to the Cell Theory. After you view the videos, attempt the following questions to test your ability to recall important contributors and tools in developing the Cell Theory.
- “Lens making in the 1600s” (4:16 min) by Corning Museum of Glass (2016).
- “Leeuwenhoek’s microscope” (5:47 min) by LabXchange (2021).
- Text background by University of California Museum of Paleontology ([date unknown]).
Answer the following questions pertaining to the videos and text reading:
- Which statement(s) is(are) true about van Leeuwenhoek’s lens making activities in the 1600s?
- He melted plastic over a flame and polished it with sandpaper.
- He ground and polished tiny pieces of glass and fine powder that were glued onto a stick.
- He also used a secret method of glass blowing a thin-walled glass tube over a flame.
- His surviving lenses are double convex, ground and polished on both sides.
Show/Hide answer.
b, c and d
- Who proposed that plant embryos arose from a single cell?
- Matthias Schleiden
- Theodor Schwann
- Robert Hooke
- Louis Pasteur
Show/Hide answer.
a. Matthias Schleiden
- Which microscope was used by Antonie van Leeuwenhoek?
- Single-lens
- Confocal microscope
- Double-lens
- Compound microscope
Show/Hide answer.
a. Single-lens
- Which of the following individuals did not contribute to the establishment of cell theory?
- Girolamo Fracastoro
- Matthias Schleiden
- Robert Remak
- Robert Hooke
Show/Hide answer.
a. Girolamo Fracastoro did not contribute to the establishment of cell theory.
Learning Activity: How Science Builds: Scientific Discovery Isn’t as Simple as One Good Experiment
This activity illuminates the twists and turns that came together to build the foundations of Cell Biology.
- Read (or re-read) the ‘Eye on Ethics: Science and Plagiarism’ excerpt.
- Watch the video Cell theory | Structure of a cell | Biology | Khan Academy (8:00 min) by Khan Academy (2015).
- Watch the video “The weird and wonderful history of cell theory” (6:00 min) by Lauren Royal-Woods at TEDEd (2012).
- Answer the following questions:
- Can you describe how science builds using examples provided about the development of specific aspects of the cell theory in these videos?
- Why do you think it was so difficult to develop the cell theory?
Basic Properties of Cells
Section Learning Objectives
By the end of this section, you will be able to:
- Describe the structural and functional aspects that are common to all cells, from organisms as different as bacteria and human beings.
- Describe how cells that are structurally diverse perform the same functions.
- Identify and describe structures and organelles unique to eukaryotic cells.
- Compare and contrast similar structures found in prokaryotic and eukaryotic cells.
Introduction
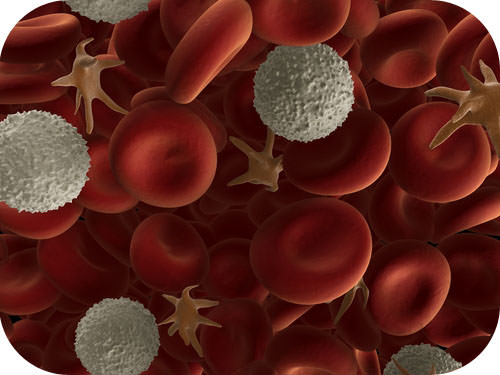
What is the same between a bacterial cell and one of your cells? The cell theory states that the cell is the fundamental unit of life. As the image of human blood in Figure 4 shows, cells come in different shapes and sizes even within one tissue, such as blood. The shapes and sizes directly influence the function of the cell. Yet, all cells—from cells in the smallest bacteria to those in the largest whale—perform some similar functions, so they do have parts in common. How did all known organisms come to have such similar structural parts and functions? The similarities show that all life on Earth has a common evolutionary history as will be discussed soon.
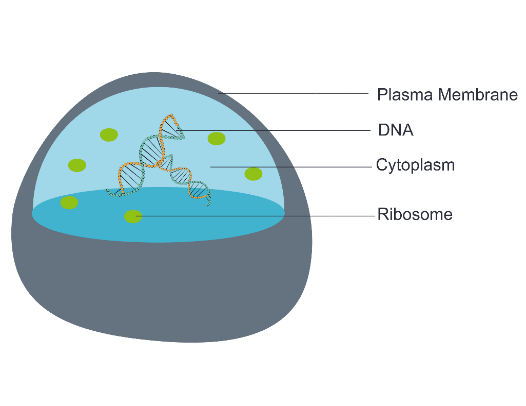
All cells (bacterial, archaeal, eukaryotic) share the following four common structural and functional components (Figure 5):
-
- The plasma membrane (a type of cell membrane) is a thin coat of lipids and proteins that surrounds a cell. It forms the physical boundary between the cell and its environment. You can think of it as the “skin” of the cell.
- Cytoplasm refers to all of the cellular material inside of the plasma membrane. Cytoplasm is made up of a jelly-like substance called cytosol, and it contains other cell structures, such as ribosomes. Cytoskeletal filaments made of proteins, including actin and tubulin, are found in the cytosol of most cells.
- Ribosomes are the structures in the cytoplasm in which proteins are made. Although all ribosomes are composed of both RNA and protein, there are some distinct differences between those found in bacteria/archaea and those found in eukaryotes, particularly in terms of size and location.
- DNA (Deoxyribonucleic acid) is the genetic material of the cell, the instructions for the cell’s abilities and characteristics. DNA is a nucleic acid found in most cells. It contains the genetic instructions that cells need to make RNA and proteins. A complete set of genes, referred to as a genome, is localized in an irregularly shaped region known as the nucleoid in bacterial and archaeal cells, but it is enclosed in a membrane-bound nucleus in eukaryotic cells. An exception is mature erythrocytes (red blood cells) that do not contain a nucleus to give more room for hemoglobin.
In addition to the common structural components listed above, all living cells have the common characteristics listed below. As you progress through this course, each of these characteristics will be considered at different levels and in different contexts.
Complexity and organization — Each cell type has a consistent structure and composition of macromolecules to maximize its specialized function
- Thermal energy from the environment causes many molecules to exhibit a random motion that is used in subsequent highly organized and directed events.
Metabolism — the web of all enzyme-catalyzed reactions in a cell or organism
- Enzymes increase the rate of chemical reactions that are required for cells to maintain the characteristics of life.
- Metabolism is required to convert food into energy for cellular processes, to convert food/fuel into macromolecules and to eliminate wastes.
Energy — the living cells of every organism constantly acquire and use energy
- Just as living things must continually consume or manufacture food to replenish their energy supplies, cells must continually produce more energy to replenish that used by the many energy-requiring chemical reactions that constantly take place.
- All cells use adenosine 5′-triphosphate (ATP) as their source of intracellular chemical energy.
- All cells carry out glycolysis to convert the energy in organic molecules into ATP.
- All cells use a membrane system to manufacture ATP.
Response to stimuli — living cells can respond to and interact with their environment
- The response of a single-celled organism like an amoeba is obvious as it moves toward nutrients in its immediate vicinity or away from an object in its path.
- In multi-cellular organisms, cells possess membrane receptors that mediate specific responses to soluble substances in their environment or present in the membranes of other cells. The responding cell may change its metabolic activity, move to another location or even commit suicide.
Mechanical activity — cells are involved in numerous mechanical activities
- Cells assemble and disassemble structures.
- Cells transport substances and organelles around, often with the help of motor proteins.
Self regulation — cells carry out a series of ordered reactions that are self-adjusted to maintain a complex, highly ordered state
- Cells use backup mechanisms to monitor and correct processes such as division, growth and differentiation.
Genetic program — cells contain molecules of DNA and a genetic code to use it
- Cells are built from and function based on information supplied in the form of genes, which are made of DNA.
- Genes supply the information to direct the formation of cellular structures, for running cellular activities, for making more of themselves and for facilitating evolution.
Reproduction — living cells give rise to other cells either sexually or asexually
- A mother cell divides into two daughter cells that contain an equal share of genetic information.
- The daughter cells do not always have an equal volume.
Cell evolution — all cells arose from a single, common ancestor present three billion years ago
- Evidence exists from the presence of common structures in all living cells, including a common genetic code, a cytoplasm, a plasma membrane and ribosomes.
- Cells continue to evolve as evidenced by the evolution of bacterial drug resistance.
Two Main Types of Cells
The evolution of life on Earth over the past 4 billion years has resulted in a huge variety of species. For more than 2,000 years, humans have been trying to classify the great diversity of life. The science of classifying organisms is called taxonomy. Classification is an important step in understanding the present diversity and past evolutionary history of life on Earth. All modern classification systems have their roots in the Linnaean classification system. It was developed by Swedish botanist Carolus Linnaeus in the 1700s. The Linnaean system of classification consists of a hierarchy of groupings, called taxa (singular, taxon). Taxa range from the kingdom to the species. The kingdom is the largest and most inclusive grouping. It consists of organisms that share just a few basic similarities. Examples are the plant and animal kingdoms. The species is the smallest and most exclusive grouping. It consists of organisms that are similar enough to produce fertile offspring together. Closely related species are grouped together in a genus.
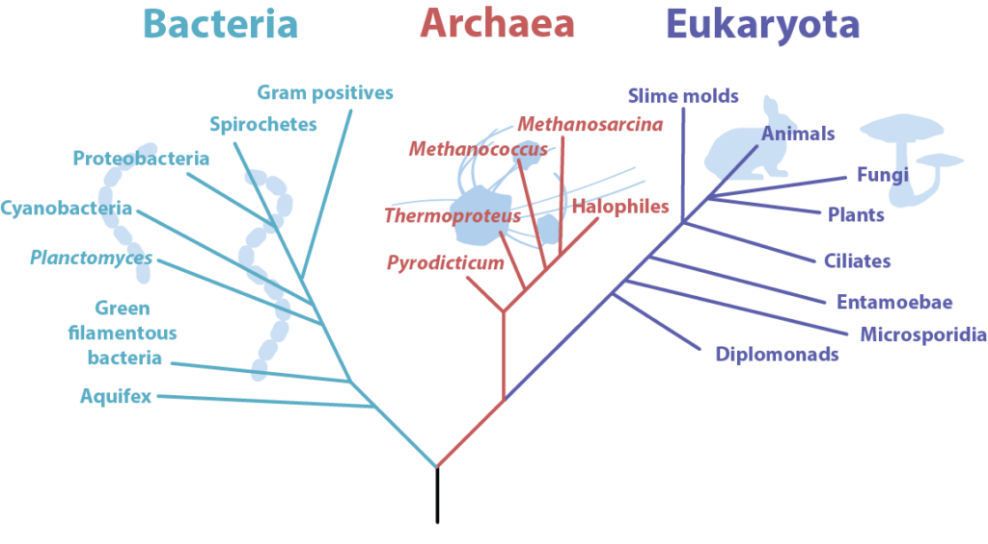
A major change to the Linnaean system was the addition of a new taxon called the domain. A domain is a taxon that is larger and more inclusive than the kingdom. Most biologists agree there are three domains of life on Earth: Bacteria, Archaea, and Eukaryota (Figure 6). Both Bacteria and Archaea consist of single-celled prokaryotes. Eukaryota consists of all eukaryotes, from single-celled protists to humans. This domain includes the Animalia (animals), Plantae (plants), Fungi (fungi), and Protista (protists) kingdoms (Figure 6).
General Comparison of Cell Structures — Prokaryotes
Although all cells share the four common structural and functional components as described above, prokaryotes differ from eukaryotic cells in several ways. The crux of their key difference can be deduced from their names: “karyose” is a Greek word meaning “nut” or “centre,” a reference to the nucleus of a cell. “Pro” means “before,” while “eu” means “true,” indicating that prokaryotes lack a nucleus (“before a nucleus”) while eukaryotes have a true nucleus.
The structures inside a cell are analogous to the organs inside a human body, with unique structures suited to specific functions. Some of the structures found in prokaryotic cells are similar to those found in some eukaryotic cells; others are unique to prokaryotes. Although there are some exceptions, eukaryotic cells tend to be larger than prokaryotic cells. The larger size of eukaryotic cells dictates the need to compartmentalize various chemical processes within different areas of the cell, using complex membrane-bound organelles. In contrast, prokaryotic cells generally lack membrane-bound organelles; however, they often contain inclusions and microcompartments that compartmentalize their cytoplasm. The structures typically associated with prokaryotic cells are illustrated in Figure 7 and summarized in Table 1.
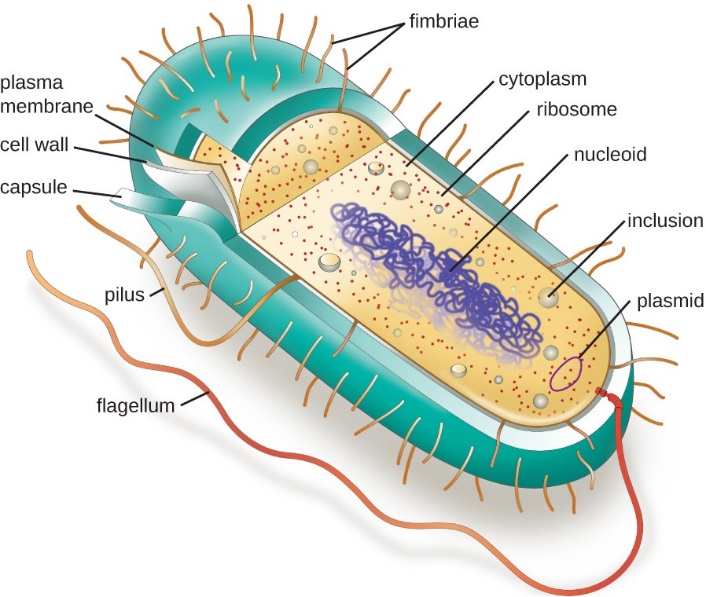
| Table 1: Prokaryotic Cell Structures. (Wakim and Grewal/Human Biology/LibreTexts) CK-12 License |
|
| Cell Structure | Description |
| Pili | Small projections outside of the cell; aid in attachment and reproduction |
| Flagellum | Long projection(s) outside of the cell in some bacteria; aids in the motility |
| Capsule | A thick protective layer outside the cell wall of some bacteria |
| Cell wall | Outer layer of bacterial cells; more chemically complex than eukaryotic cell walls |
| Plasma membrane | Phospholipid bilayer marking the outside of the cytoplasm |
| Cytoplasm | The fluid portion of the cell |
| Ribosome | Involved in protein synthesis |
| Nucleoid | Circular DNA found in the cytoplasm |
| Plasmid | Small loops of DNA found in some bacteria |
More recently, microbiologists are resisting the term prokaryote because it lumps both bacteria and the subsequently discovered archaea in the same category. Both cells are prokaryotic because they lack a nucleus and other organelles (such as mitochondria, Golgi apparatus, endoplasmic reticulum, etc.), but they aren’t closely related genetically. Therefore, to honour these differences we will refer to the groups as the archaea, the bacteria, and the eukaryotes.
Self-Check
True or False: All prokaryotes possess flagella, pili, fimbriae, and capsules.
Show/Hide answer
False
Eukaryotes
Eukaryotic organisms include protozoans, algae, fungi, plants, and animals. Some eukaryotic cells are independent, single-celled microorganisms, whereas others are part of multicellular organisms. The cells of eukaryotic organisms have several distinguishing characteristics. Above all, eukaryotic cells are defined by the presence of a nucleus surrounded by a complex nuclear membrane. Also, eukaryotic cells are characterized by the presence of membrane-bound organelles in the cytoplasm. The word “organelle” means “little organ,” each of which has specialized cellular functions, just as your body’s organs have specialized functions.
Organelles such as mitochondria, the endoplasmic reticulum (ER), Golgi apparatus, lysosomes, and peroxisomes are held in place by the cytoskeleton, an internal network that supports transport of intracellular components and helps maintain cell shape (Figure 8). The genome of eukaryotic cells is packaged in multiple, rod-shaped chromosomes as opposed to the single, circular-shaped chromosome that characterizes most prokaryotic cells.
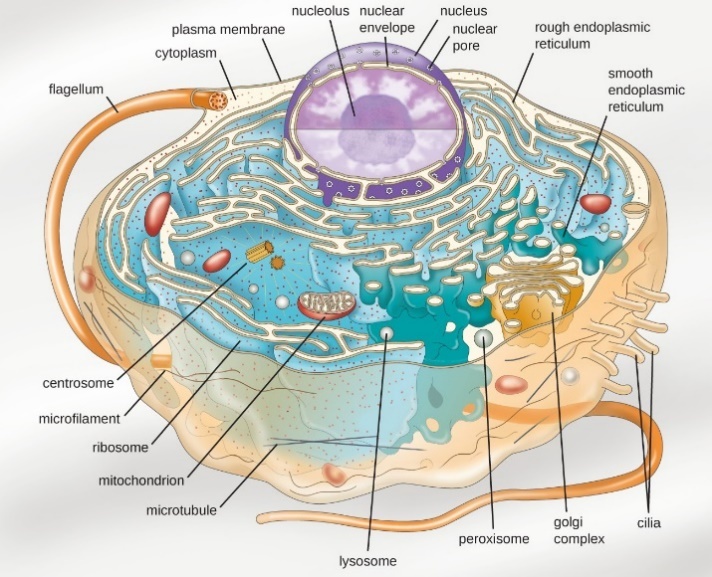
Have you ever heard the phrase “form follows function?” It’s a philosophy that many industries follow. In architecture, this means buildings should be constructed to support the activities that will be carried out inside them. For example, a skyscraper should include several elevator banks, and a hospital should have its emergency room easily accessible.
Our natural world also uses the principle of form follows function, especially in cell biology, and this will become clear as we explore the structure and function of eukaryotic cells. Table 2 compares the characteristics of eukaryotic cell structures with those of bacteria and archaea.
| Table 2: Comparing Cell Structures in Bacteria, Archaea and Eukaryotes. (Adapted from Parker et al. 2016/ Microbiology/ OpenStax) CC BY 4.0 |
|||
| Cell Structure | Prokaryotes | Eukaryotes | |
| Bacteria | Archaea | ||
| Size | ~0.5–1 μm | ~0.5–1 μm | ~5–20 μm |
| Surface area-to-volume ratio | High | High | Low |
| Nucleus | No | No | Yes |
| Genome characteristics | · Single chromosome · Circular · Haploid · Lacks histones |
· Single chromosome · Circular · Haploid · Contains histones |
· Multiple chromosomes · Linear · Haploid or diploid · Contains histones |
| Cell division | Binary fission | Binary fission | · Mitosis · Meiosis |
| Membrane lipid composition | · Ester-linked straight-chain fatty acids · Bilayer |
· Ether-linked branched isoprenoids · Monolayer or bilayer |
· Ester-linked straight-chain fatty acids · Cholesterol, sterols · Bilayer |
| Cell wall composition | · Peptidoglycan · None |
· Pseudopeptidoglycan · Glycopeptide · Polysaccharide · Protein (S-layer) · None |
· Cellulose (plants, some algae) · Chitin (molluscs, insects, crustaceans, fungi) · Silica (some algae) · Most others lack cell walls |
| Motility structures | Rigid spiral flagella composed of flagellin | Rigid spiral flagella composed of archaeal flagellins | Flexible flagella and cilia composed of microtubules |
| Membrane-bound organelles | No | No | Yes |
| Endomembrane system | No | No | Yes (ER, Golgi, lysosomes) |
| Ribosomes | 70S | 70S | · 80S (cytoplasm, rough ER) · 70S (mitochondria, chloroplasts) |
Cell Morphology
Individual cells of a particular prokaryotic organism are typically similar in shape, or cell morphology. The shape dictates how that cell will grow, reproduce, obtain nutrients, move, and it’s important to the cell to maintain that shape to function properly. Cell morphology can be used as a characteristic to assist in identifying particular microbes, but it’s important to note that cells with the same morphology are not necessarily related. Although thousands of prokaryotic organisms have been identified, only a handful of cell morphologies are commonly seen microscopically. Figure 9 names and illustrates cell morphologies commonly found in prokaryotic cells.
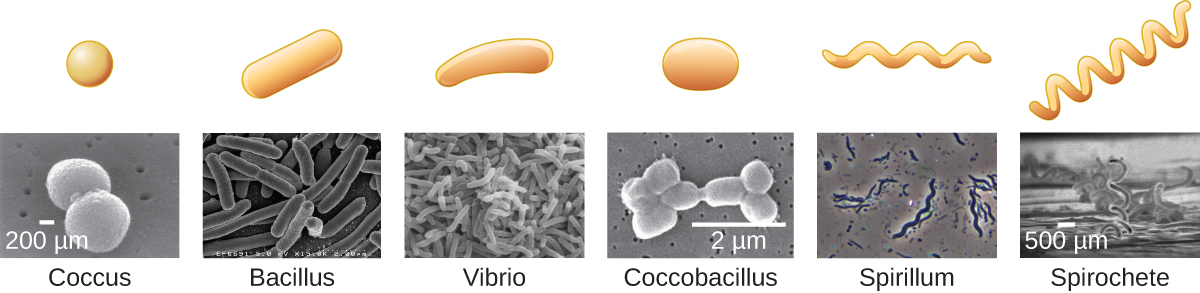
There are additional shapes seen for bacteria, and an even wider array for the archaea, which have even been found as star or square shapes. Eukaryotic microbes also tend to exhibit a wide array of shapes, particularly the ones that lack a cell wall such as the protozoa.
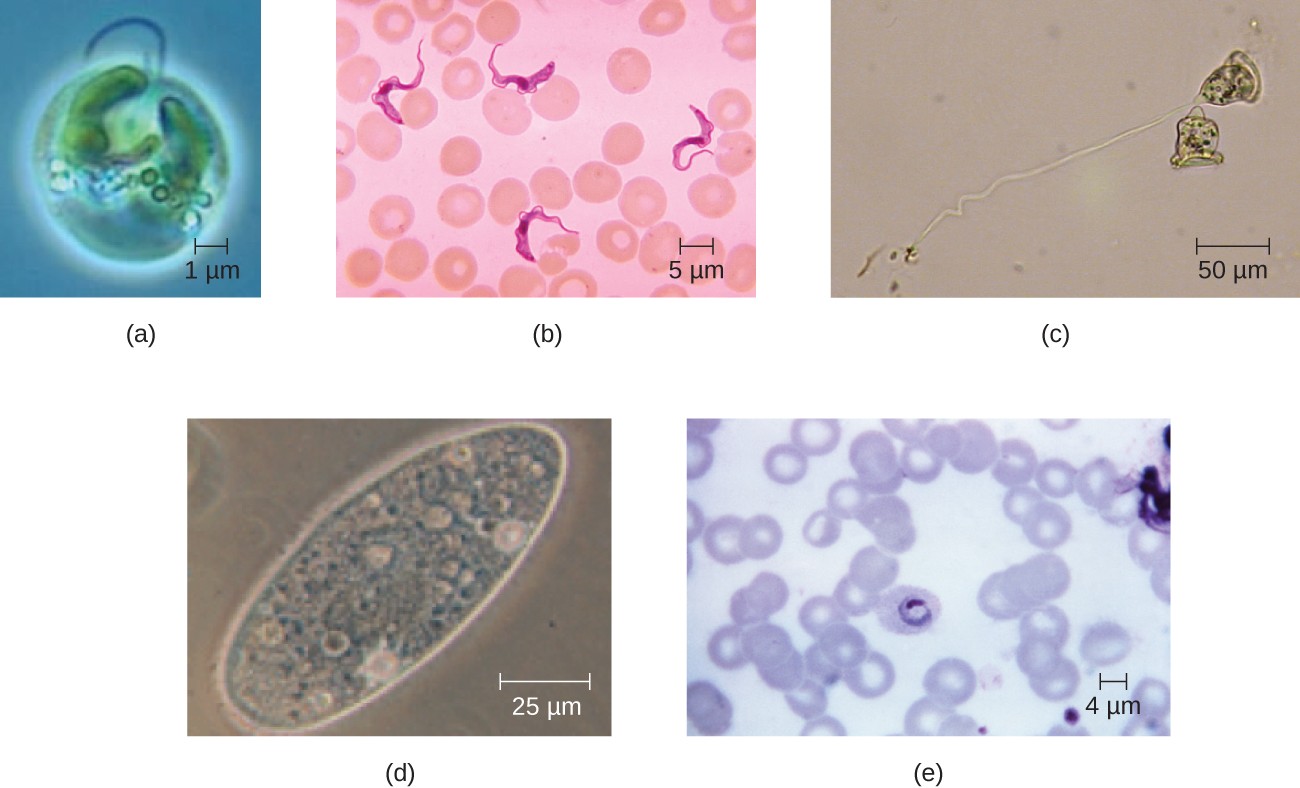
Eukaryotic cells also display a wide variety of different cell morphologies. Possible shapes include spheroid, ovoid, cuboidal, cylindrical, flat, lenticular, fusiform, discoidal, crescent, ring stellate, and polygonal (Figure 10). Some eukaryotic cells are irregular in shape, and some are capable of changing shape. The shape of a particular type of eukaryotic cell may be influenced by factors such as its primary function, the organization of its cytoskeleton, the viscosity of its cytoplasm, the rigidity of its cell membrane or cell wall (if it has one), and the physical pressure exerted on it by the surrounding environment and/or adjoining cells.
Ribosomes
All cellular life synthesizes proteins, and organisms in all three domains of life possess ribosomes, structures responsible for protein synthesis. However, ribosomes in each of the three domains are structurally different. Ribosomes are evolutionarily conserved protein synthesizing machines found in all cells (see Table 2). Because protein synthesis is essential for all cells, ribosomes are found in practically every cell, although they are smaller in prokaryotic cells. Ribosomes are particularly abundant in immature red blood cells for the synthesis of hemoglobin, which functions in the transport of oxygen throughout the body.
Ribosomes consist of a large and a small subunit, each made up of multiple proteins and one or more molecules of ribosomal RNA (rRNA). Electron microscopy has shown that ribosomes consist of two subunits, aptly called large and small (Figure 11). Ribosomes receive their “instructions” for protein synthesis from the nucleus, where the DNA is transcribed into messenger RNA (mRNA). The mRNA exits through nuclear pores and travels to the ribosomes. To translate the mRNA code into an amino acid sequence, transfer RNA (tRNA) with appropriate anticodons, complementary nucleotide sequence to mRNA, and bind to the ribosome-mRNA complex. Each tRNA brings a specific amino acid that link together to form the polypeptide chain of a protein.
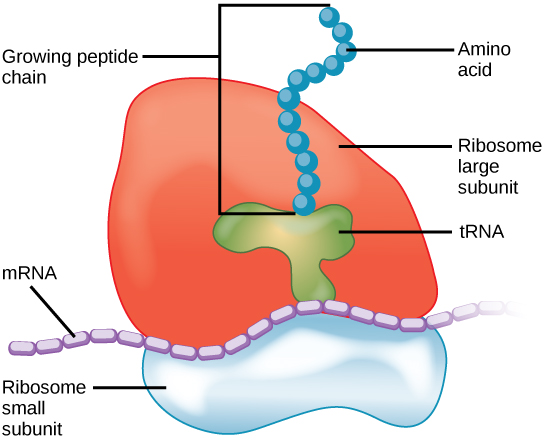
Eukaryotic and prokaryotic ribosomes differ in the number of RNA and proteins in their large and small subunits and, thus, in their overall size. Isolated ribosomes centrifuged in a sucrose density gradient move at a rate based on their size (Figure 12).
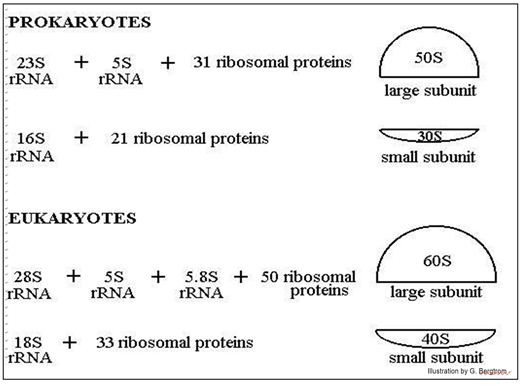
The position of ribosomal subunits in the gradient is represented by an S value, after Svedberg, who first used sucrose density gradients to separate macromolecules and particles by size. Basically, it is a sedimentation coefficient that measures how fast a particle moves to the bottom of a solution. Note that the ribosomal RNAs themselves also separate on sucrose density gradients by size, hence their different S values. Ribosomes found in eukaryotic organelles such as mitochondria or chloroplasts have 70S ribosomes—the same size as prokaryotic ribosomes. However, non-organelle-associated ribosomes in eukaryotic cells are 80S ribosomes, composed of a 40S small subunit and a 60S large subunit. In terms of size and composition, this makes them distinct from the ribosomes of prokaryotic cells.
Although they are the same size, bacterial and archaeal ribosomes have different proteins and rRNA molecules, and the archaeal versions are more like their eukaryotic counterparts than those found in bacteria. In fact, the rRNA nucleotide differences provided scientists with conclusive evidence to argue that archaea deserved a domain separate from the bacteria. In addition, archaeal ribosomes have a different shape than bacterial ribosomes, with proteins unique to archaea. This provides them with resistance to antibiotics that inhibit ribosomal function in bacteria.
The differences between eukaryotic and prokaryotic ribosomes are clinically relevant because certain antibiotic drugs are designed to target one or the other. For example, cycloheximide targets eukaryotic action, whereas chloramphenicol targets prokaryotic ribosomes (Barnhill et al. 2012). Generally, since human cells are eukaryotic, they are unharmed by antibiotics that destroy the prokaryotic ribosomes in bacteria. However, sometimes negative side effects may occur because mitochondria in human cells contain prokaryotic ribosomes.
Self-Check
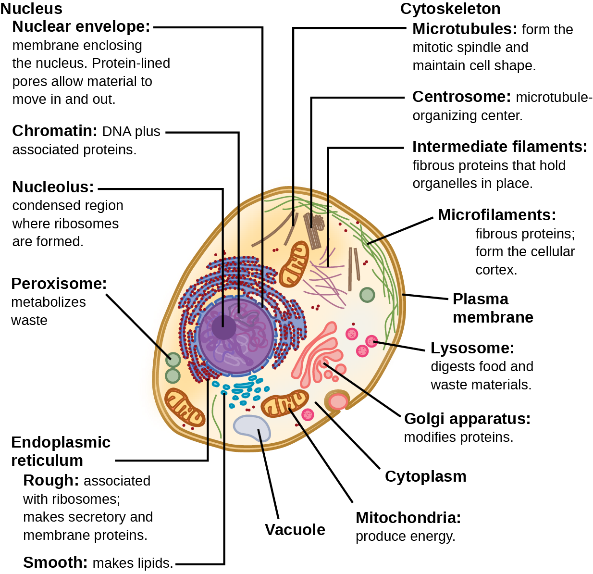
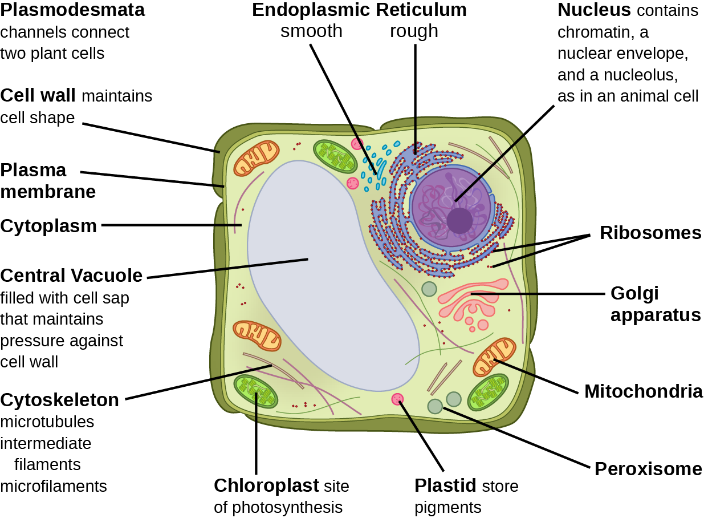
While all eukaryotic cells contain the previously mentioned organelles and structures, animal and plant cells have some striking differences (compare Figure 13 and Figure 14; Table 3).
The following table provides the components of prokaryotic and eukaryotic cells and their respective functions:
| Table 3: Components of Prokaryotic and Eukaryotic Cells and Their Functions. (Adapted from Fowler et al. 2013) CC BY 4.0 | |||
| Cell Component and Function | Present in Prokaryotes? | Present in Animal Cells? | Present in Plant Cells? |
|---|---|---|---|
| Plasma membrane Separates the cell from the external environment; controls passage of organic molecules, ions, water, oxygen, and wastes into and out of the cell |
Yes | Yes | Yes |
| Cytoplasm Provides structure to cell; site of many metabolic reactions; medium in which organelles are found |
Yes | Yes | Yes |
| Nucleoid Location of DNA |
Yes | No | No |
| Nucleus Cell organelle that houses DNA and directs synthesis of ribosomes and proteins |
No | Yes | Yes |
| Ribosomes Protein synthesis |
Yes | Yes | Yes |
| Mitochondria ATP production, cellular respiration |
No | Yes | Yes |
| Peroxisomes Oxidizes and breaks down fatty acids and amino acids, detoxifies poisons |
No | Yes | Yes |
| Vesicles and vacuoles Storage and transport; digestive function in plant cells |
No | Yes | Yes |
| Centrosome Unspecified role in cell division in animal cells; organizing centre of microtubules in animal cells |
No | Yes | No |
| Lysosomes Digestion of macromolecules; recycling of worn-out organelles |
No | Yes | No |
| Cell wall Protection, structural support and maintenance of cell shape |
Yes, primarily peptidoglycan in bacteria but not Archaea | No | Yes, primarily cellulose |
| Chloroplasts Photosynthesis |
No | No | Yes |
| Endoplasmic reticulum Modifies proteins and synthesizes lipids |
No | Yes | Yes |
| Golgi apparatus Modifies, sorts, tags, packages, and distributes lipids and proteins |
No | Yes | Yes |
| Cytoskeleton Maintains cell’s shape, secures organelles in specific positions, allows cytoplasm and vesicles to move within the cell, and enables unicellular organisms to move independently |
Yes | Yes | Yes |
| Flagella Cellular locomotion |
Some | Some | No, except for some plant sperm |
| Cilia Cellular locomotion, movement of particles along the extracellular surface of the plasma membrane, and filtration |
No | Some | No |
Learning Activity: Parts of a Cell
- Watch the video “Parts of a cell” (21 min) by Khan Academy (2010) to review the parts of a cell.
- Answer the following questions:
You have been asked to characterize the organisms in a sample you obtained from a recent field trip by light and electron microscopy.
- How would you convince yourself and others that your organisms are ‘alive’?
- How would you determine whether your organisms are prokaryotes, eukaryotes and/or neither?
Learning Activity: Cell Comparisons
LabXchange, a digital interactive classroom site created at Harvard University, provides a variety of lab simulations, videos and other content to help support active science learning. At various points in this course, you will be asked to complete activities at this site. You are not required to create an account to complete the activities in this course, but you may wish to explore this option on your own.
- Complete the interactive exercise “Comparing Four Cells: Animal, Plant, Bacteria, Yeast” by SERP Institute (2021) on LabXChange, where you will compare four different cells: animal, plant, bacteria and yeast. When you go to the link, you will be asked to “start the interactive,” where you can view the different kinds of cells.
- Click on each cellular feature and compare its presence between prokaryotic and various types of eukaryotic cells.
Learning Activity: Cell Structure and Function
- Go to the simulation activity “Cell Structure” by Biological Simulations (2022) on LabXchange, where you can practice identifying cell parts by name or function.
- This WISC-Online interactive website can be used as a review throughout this course. In the activity “A typical animal cell” (Heins [date unknown]), you can practice identifying cell parts by name or function.
Origin of Eukaryotic Cells
Section Learning Objectives
By the end of this section, you will be able to:
- Describe what scientists know about the origins of eukaryotes based on the last common ancestor.
- Define endosymbiosis.
- Explain the key points of endosymbiotic theory and cite the evidence that supports this concept.
- Explain what “slug power” means and how it relates to endosymbiosis and the evolution of cells.
Endosymbiosis and the Evolution of Eukaryotes
Organisms are classified into three domains: Archaea, Bacteria, and Eukarya. The first two lineages comprise all prokaryotic cells, and the third contains all eukaryotes. A very sparse fossil record prevents us from determining what the first members of each of these lineages looked like, so all the events that led to the last common ancestor of extant (living) eukaryotes may remain unknown. However, the comparative biology of extant (surviving) organisms and the limited fossil record provide some insight into the evolution of Eukarya.
The earliest fossils found appear to be those of domain Bacteria, most likely cyanobacteria. They are about 3.5 to 3.8 billion years old and are recognizable because of their relatively complex structure and, for prokaryotes, relatively large cells. Most other prokaryotes have small cells, 1 or 2 µm in size, and would be difficult to pick out as fossils. Fossil stromatolites suggest that at least some prokaryotes lived in interactive communities, and evidence from the structure of living eukaryotic cells suggests that similar ancestral interactions gave rise to the eukaryotes. Most living eukaryotes have cells measuring 10 µm or greater. Structures this size, which might be fossilized remains of early eukaryotes, appear in the geological record in deposits dating to about 2.1 billion years ago.
Data from these fossils and the study of living genomes have led comparative biologists to conclude that living eukaryotes are all descendants of a single common ancestor. Mapping the characteristics found in all major groups of eukaryotes reveals that the common characteristics are present in at least some of the members of each major lineage or during some part of their life cycle and, therefore, must have been present in what is known as the last universal common ancestor (LUCA). LUCA is the most recent common ancestor of all current life on Earth, estimated to have lived some 3.5 to 3.8 billion years ago. LUCA is not thought to be the first life on Earth, but rather the only type of organism of its time to still have living descendants.
Before discussing the origins of eukaryotes in more detail, it is first important to understand that all extant (living) eukaryotes are likely the descendants of a chimera-like organism that was a composite of a host cell and the cell(s) of an alpha-proteobacterium that “took up residence” inside it. This major theme in the origin of eukaryotes is endosymbiosis, one cell engulfing another such that the engulfed cell survives and both cells benefit. Over many generations, a symbiotic relationship can result in two organisms that depend on each other so much that neither can survive on its own. Endosymbiotic events likely contributed to the origin of the last common ancestor of today’s eukaryotes and to later diversification in certain lineages of eukaryotes. Before explaining this further, first consider metabolism in prokaryotes.
Self-Check
Why is it agreed that prokaryotes arose before eukaryotes?
Show/Hide answer
Fossil record showing prokaryotes at 3.5 to 3.8 bya and eukaryotes not until 1 billion years later.
Prokaryotic Metabolism
Many important metabolic processes arose in prokaryotes; however, some, such as nitrogen fixation, are never found in eukaryotes. All major lineages of eukaryotes have aerobic respiration localized in the mitochondria. Aerobic respiration is also found in many lineages of prokaryotes but not in all of them, and a great deal of evidence suggests that neither these anaerobic prokaryotes nor their ancestors ever carried out aerobic respiration.
While today’s atmosphere is about 20 percent molecular oxygen (O2), geological evidence shows it previously lacked O2. Without oxygen, living things would not use aerobic respiration; instead, they would have relied on anaerobic respiration or fermentation. At some point before, about 3.8 billion years ago, some prokaryotes began using energy from sunlight to power anabolic processes that reduce carbon dioxide to form organic compounds. That is, they evolved the ability to photosynthesize. Hydrogen, derived from various sources, was “captured” using light-powered reactions to reduce fixed carbon dioxide in the Calvin cycle. The group of Gram-negative bacteria that gave rise to cyanobacteria used water as the hydrogen source and released O2 as a “waste” product.
Eventually, the amount of photosynthetic oxygen built up in some environments to levels that posed a risk to living organisms since it can damage many organic compounds. Various metabolic processes evolved that protected organisms from oxygen, one of which, aerobic respiration, also generated high levels of ATP. It became widely present among prokaryotes, including in a free-living group now called alpha-proteobacteria. Organisms that did not acquire aerobic respiration had to remain in oxygen-free environments. In the past, oxygen-rich environments likely only occurred around places where cyanobacteria were abundant and active; however, about 2 billion years ago, geological evidence shows that the atmosphere began to build up higher oxygen concentrations. Oxygen levels similar to today’s levels only arose within the last 700 million years.
Recall that the first fossils believed to be eukaryotes date to about two billion years old, so they seem to have evolved and diversified rapidly as oxygen levels increased. Also, recall that all extant eukaryotes descended from an ancestor with mitochondria. Light microscopists first observed these organelles in the late 1800s, where they appeared as somewhat worm-shaped structures that seemed to move around in the cell. Some early observers suggested they might be bacteria living inside host cells, but these hypotheses remained unknown or rejected in most scientific communities, as described below.
Development of the Endosymbiotic Theory
As scientists progressed toward understanding the role of cells in plant and animal tissues, others examined the structures within the cells themselves. In 1831, Scottish botanist Robert Brown (1773–1858) was the first to describe nuclei, which he observed in plant cells. Then, in the early 1880s, German botanist Andreas Schimper (1856–1901) was the first to describe the chloroplasts of plant cells, identifying their role in starch formation during photosynthesis and noting that they divided independently of the nucleus.
Based upon the chloroplasts’ ability to reproduce independently, Russian botanist Konstantin Mereschkowski (1855–1921) suggested in 1905 that chloroplasts may have originated from ancestral photosynthetic bacteria living symbiotically inside a eukaryotic cell. He proposed a similar origin for the nucleus of plant cells. This proposal was the first articulation of the endosymbiotic hypothesis, and it would explain how eukaryotic cells evolved from ancestral bacteria.
American anatomist Ivan Wallin (1883–1969) furthered Mereschkowski’s endosymbiotic hypothesis by experimentally examining the similarities between mitochondria, chloroplasts, and bacteria—in other words, to put the endosymbiotic hypothesis to the test using objective investigation. Wallin published a series of papers in the 1920s supporting the endosymbiotic hypothesis, including a 1926 publication co-authored with Mereschkowski. Wallin claimed he could culture mitochondria outside of their eukaryotic host cells. Many scientists dismissed his cultures of mitochondria as resulting from bacterial contamination. Modern genome sequencing work supports the dissenting scientists by showing that much of the genome of mitochondria had been transferred to the host cell’s nucleus, preventing the mitochondria from living by themselves (Embley and Martin 2006; Berg and Kurland 2000).
The scientific community largely ignored Wallin’s ideas regarding the endosymbiotic hypothesis for the next 50 years because scientists were unaware that these organelles contained their own DNA. However, the discovery of mitochondrial and chloroplast DNA in the 1960s resurrected the endosymbiotic hypothesis. Lynn Margulis (1938–2011), an American geneticist, published her ideas regarding the endosymbiotic hypothesis of the origins of mitochondria and chloroplasts in 1967 (Sagan 1967). In the decade leading up to her publication, advances in microscopy had allowed scientists to differentiate prokaryotic cells from eukaryotic cells. In her publication, Margulis reviewed the literature and argued that the eukaryotic organelles, such as mitochondria and chloroplasts, are of prokaryotic origin. She presented a growing body of microscopic, genetic, molecular biology, fossil, and geological data to support her claims.
Again, this hypothesis was not initially popular. However, mounting genetic evidence due to the advent of DNA sequencing supported the endosymbiotic theory, now defined as the theory that mitochondria and chloroplasts arose because prokaryotic cells established a symbiotic relationship within an eukaryotic host (Figure 15). The endosymbiotic theory states that eukaryotes may have been a product of one cell engulfing another, one living within another, and coevolving over time until the separate cells were no longer recognizable as such and shared genetic control of a mutualistic metabolic pathway to produce ATP.
With Margulis’ initial endosymbiotic theory gaining wide acceptance, she expanded on the theory in her 1981 book Symbiosis in Cell Evolution. In it, she explains how endosymbiosis is a major driving factor in the evolution of organisms. More recent genetic sequencing and phylogenetic analysis show that mitochondrial and chloroplast DNA are highly related to their bacterial counterparts in DNA sequence and chromosome structure. However, mitochondrial and chloroplast DNA are reduced compared to nuclear DNA because many genes have moved from the organelles into the host cell’s nucleus. Additionally, mitochondrial and chloroplast ribosomes are structurally similar to bacterial ribosomes rather than the eukaryotic ribosomes of their hosts. Last, the binary fission of these organelles strongly resembles the binary fission of bacteria, as compared with mitosis performed by eukaryotic cells. Since Margulis’ original proposal, scientists have observed several examples of bacterial endosymbionts in modern-day eukaryotic cells. Examples include the endosymbiotic bacteria found within the guts of certain insects, such as cockroaches (Douglas 2009), and photosynthetic bacteria-like organelles found in protists (Jaynes and Vernon 1982).
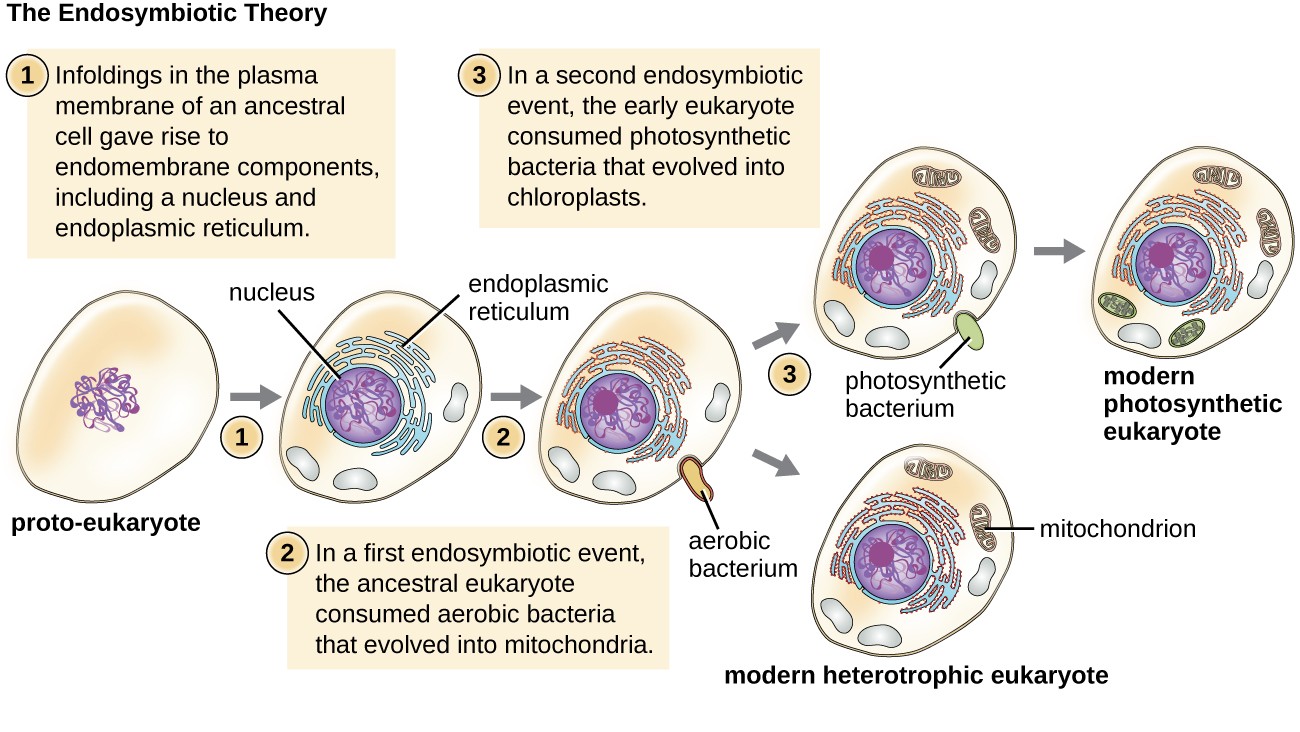
Although Margulis’s work initially was met with resistance, this basic component of this once-revolutionary hypothesis is now widely accepted, with work progressing on uncovering the steps involved in this evolutionary process and the key players involved.
Mitochondria — The Evidence
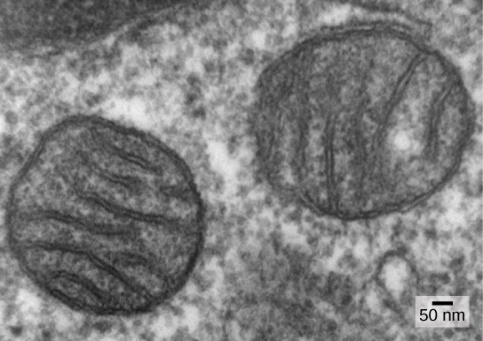
One of the major features distinguishing prokaryotes from eukaryotes is the presence of mitochondria, or their reduced derivatives, in virtually all eukaryotic cells. Eukaryotic cells may contain anywhere from one to several thousand mitochondria, depending on the cell’s level of energy consumption, in humans being most abundant in the liver and skeletal muscles. Each mitochondrion measures 1 to 10 or greater micrometers in length and exists in the cell as an organelle that can be ovoid to worm-shaped to intricately branched (Figure 16). However, although they may have originated as free-living aerobic organisms, mitochondria can no longer survive and reproduce outside the cell.
Mitochondria have several features that suggest their relationship to alpha-proteobacteria (Figure 16). Alpha-proteobacteria are a large group of bacteria that includes species symbiotic with plants, disease organisms that can infect humans via ticks, and many free-living species that use light for energy. Mitochondria have their own genomes, with a circular chromosome stabilized by attachments to the inner membrane. Mitochondria also have special ribosomes and transfer RNAs resembling these same components in prokaryotes. An intriguing feature of mitochondria is that many of them exhibit minor differences from the universal genetic code. However, many of the genes for respiratory proteins are now in the nucleus. When comparing these genes to those of other organisms, they appear to be of alpha-proteobacterial origin. In some eukaryotic groups, such genes are found in the mitochondria, whereas in other groups, they are in the nucleus. Scientists have interpreted this difference as evidence that, over evolutionary time, genes have transferred from the endosymbiont chromosome to those of the host genome. This apparent “loss” of genes by the endosymbiont is probably one explanation for why mitochondria cannot live without a host.
Another line of evidence supporting the idea that mitochondria came from endosymbiosis is the structure of the mitochondrion itself. Most mitochondria have a shape similar to alpha-proteobacteria. They have two surrounding membranes: the inner membrane is bacterial in nature, while the outer membrane is eukaryotic. This membrane characteristic is exactly what one would expect if one membrane-bound organism was engulfed into a vacuole by another membrane-bound organism. The outer mitochondrial membrane was derived from the enclosing vesicle, while the inner membrane was from the endosymbiont’s plasma membrane. The extensive mitochondrial inner membrane involves substantial infoldings, called cristae, that resemble the textured outer surface of alpha-proteobacteria. The matrix and inner membrane are rich with the enzymes necessary for aerobic respiration.
The third line of evidence comes from the production of new mitochondria. Mitochondria divide independently by a process that resembles binary fission in prokaryotes. Mitochondria arise only from previous mitochondria; they are not formed from scratch (de novo) by the eukaryotic cell. Mitochondria may fuse together and move around inside the cell by interacting with the cytoskeleton. They reproduce within their enclosing cell and are distributed with the cytoplasm when a cell divides or when two cells fuse. Therefore, although these organelles have become highly integrated into the eukaryotic cell, they still reproduce as if they were independent organisms within the cell. However, their reproduction has synchronized with the activity and division of the cell. These features all support the theory that mitochondria were once free-living prokaryotes.
Some living eukaryotes are anaerobic and cannot survive around too much oxygen. However, a few appear to lack organelles that could be mitochondria. In the 1970s into the early 1990s, many biologists suggested that some of these eukaryotes descended from ancestors whose lineages had diverged from the lineage of mitochondrion-containing eukaryotes before endosymbiosis occurred. Later findings suggested that reduced organelles are found in most, if not all, anaerobic eukaryotes and that virtually all eukaryotes appear to carry some genes in their nuclei that are of mitochondrial origin.
In addition to the aerobic generation of ATP, mitochondria have several other metabolic functions. One of these functions is to generate clusters of iron and sulfur that are important cofactors of many enzymes. Such functions are often associated with the reduced mitochondrion-derived organelles of anaerobic eukaryotes. The protist Monocercomonoides, an inhabitant of vertebrate digestive tracts, appears to be an exception; it has no mitochondria, and its genome contains neither genes derived from mitochondria nor nuclear genes related to mitochondrial maintenance. However, it is related to other protists with reduced mitochondria and probably represents an end-point in mitochondrial reduction. Although most biologists accept that the last common ancestor of eukaryotes had mitochondria, it appears that the complex relationship between mitochondria and their host cell continues to evolve.
Maternal Inheritance of Mitochondria
Many cultures of the world trace their ancestry through the father’s side. For example, children take on their father’s last name. However, there are also traditions that are matrilineal. Some Jewish traditions inherit the Jewish identity through the mother, and in multiple pre-contact Indigenous cultures, wealth and power became passed down maternally.
It may seem trivial to ask whether someone’s biological composition is exclusively paternal or maternal because both the father and the mother contribute to genetic inheritance. DNA is organized as chromosomes in the nucleus, and each pair of chromosomes is inherited from both parents, one paternal copy and one maternal copy. Interestingly, mitochondria are usually inherited maternally as the mitochondria residing in the oocyte. Thus, mitochondrial DNA is helpful for tracing someone’s maternal ancestry.
Plastids — The Evidence
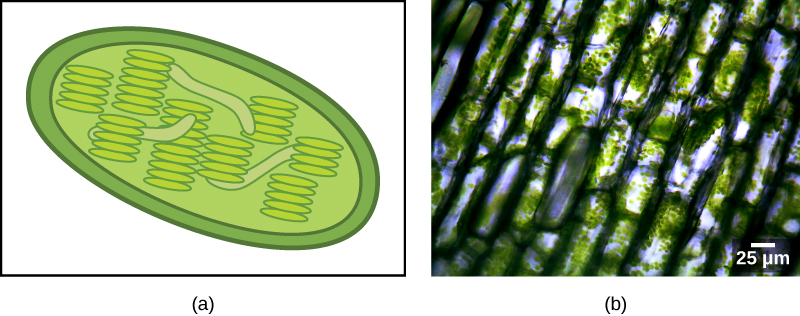
Some groups of eukaryotes are photosynthetic. Their cells contain, in addition to the standard eukaryotic organelles, another kind of organelle called a plastid. When such cells carry out photosynthesis, their plastids are rich in the pigment chlorophyll a and a range of other pigments, called accessory pigments, which are involved in harvesting energy from light. Photosynthetic plastids are called chloroplasts (Figure 17).
Like mitochondria, plastids appear to have an endosymbiotic origin. This hypothesis was also proposed and championed with the first direct evidence by Lynn Margulis. Scientists now know that plastids are from the cyanobacteria that used to live inside the cells of an ancestral aerobic heterotrophic eukaryote. Plastids of primary origin integrated into the eukaryotic cells through primary endosymbiosis, resulting in two surrounding membranes. However, the best evidence is that the acquisition of cyanobacterial endosymbionts has happened twice in the history of eukaryotes. In one case, the common ancestor of the major lineage/supergroup Archaeplastida took on a cyanobacterial endosymbiont; in the other, the ancestor of the small amoeboid rhizarian taxon, Paulinella, took on a different cyanobacterial endosymbiont. Almost all photosynthetic eukaryotes descended from the first event, and only a couple of species are derived from the other, which, in evolutionary terms, appears to be more recent.
Cyanobacteria are a group of Gram-negative bacteria with all the conventional structures of the group. However, unlike most prokaryotes, they have extensive, internal membrane-bound sacs called thylakoids. Chlorophyll is a component of these membranes, as are many of the proteins of the light reactions of photosynthesis. Cyanobacteria also have the peptidoglycan wall and lipopolysaccharide layer associated with Gram-negative bacteria.
Chloroplasts of primary endosymbiotic origin have thylakoids, a circular DNA chromosome, and ribosomes similar to cyanobacteria. As in mitochondria, each chloroplast has two surrounding membranes. Scientists believe the outer membrane is from the enclosing vacuole of the host and the inner membrane from the plasma membrane of the cyanobacterial endosymbiont. In the group of Archaeplastida called the glaucophytes and the rhizarian Paulinella, a thin peptidoglycan layer is still present between the outer and inner plastid membranes. All other plastids lack this relic of the cyanobacterial wall.
There is also, as with the case of mitochondria, strong evidence that many of the endosymbiont genes transferred to the nucleus. Plastids, like mitochondria, cannot live independently outside the host. In addition, like mitochondria, plastids are derived from the division of other plastids and are never built from scratch. Researchers have suggested that the endosymbiotic event that led to Archaeplastida occurred 1 to 1.5 billion years ago, at least five hundred million years after the fossil record suggests that eukaryotes were present.
Not all plastids in eukaryotes are derived directly from primary endosymbiosis. Some of the major groups of algae became photosynthetic by secondary endosymbiosis, that is, by taking in either green algae or red algae (both from Archaeplastida) as endosymbionts (Figure 18). Numerous microscopic and genetic studies have supported this conclusion. Secondary plastids have three or more surrounding membranes, and some secondary plastids even have clear remnants of the nucleus (nucleomorphs) of endosymbiotic algae. There are even cases where tertiary or higher-order endosymbiotic events are the best explanations for the features of some eukaryotic plastids.
Self-Check
Why is it agreed that prokaryotes gave rise to eukaryotes?
Show/Hide answer
These two types of cells must be related because they share many similar traits, such as genetic codes, enzymes, metabolic pathways and plasma membranes.
Secondary Endosymbiosis in Chlorarachniophytes
Endosymbiosis involves one cell engulfing another to produce, over time, a coevolved relationship in which neither cell can survive alone. For instance, red and green algae chloroplasts formed when an ancestral prokaryote engulfed a photosynthetic cyanobacterium.
This evidence suggests the possibility that an ancestral cell (already containing a photosynthetic endosymbiont) was engulfed by another eukaryote cell, resulting in a secondary endosymbiosis. Molecular and morphological evidence suggests chlorarachniophyte protists formed from a secondary endosymbiotic event. Chlorarachniophytes are rare algae indigenous to tropical seas and sand. They are part of the Rhizarian supergroup. Chlorarachniophytes are reticulose amoebae, extending thin cytoplasmic strands that interconnect them with other chlorarachniophytes in a cytoplasmic network. Scientists believe these protists originated from a eukaryote engulfing a green alga, the latter of which had previously established an endosymbiotic relationship with a photosynthetic cyanobacterium (Figure 19).
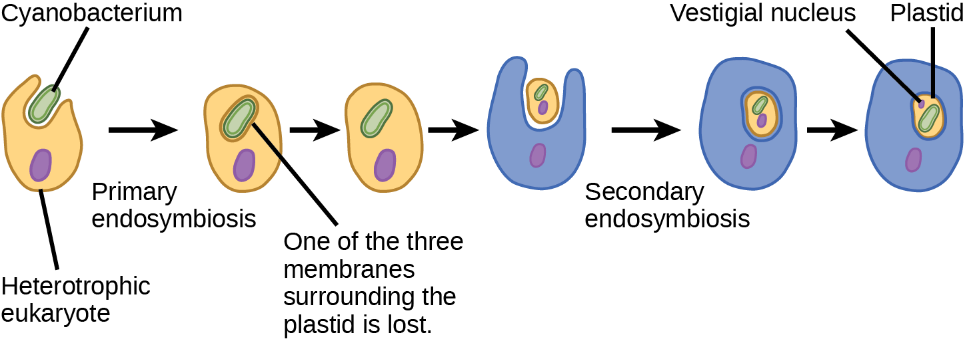
Several lines of evidence support that chlorarachniophytes evolved from secondary endosymbiosis. The chloroplasts within the green algal endosymbionts are still capable of photosynthesis, making chlorarachniophytes photosynthetic. The green algal endosymbiont also exhibits a vestigial nucleus. In fact, chlorarachniophytes appear to be the products of an evolutionarily recent secondary endosymbiotic event. Chlorarachniophytes plastids have four surrounding membranes:
- The first two correspond to the inner and outer membranes of the photosynthetic cyanobacterium.
- The third corresponds to the plasma membrane of the green alga.
- The fourth corresponds to the vacuole that surrounds the green alga when it became engulfed by the chlorarachniophyte ancestor.
In other lineages involving secondary endosymbiosis, there are only three identifiable membranes around plastids. This missing membrane is currently interpreted as a sequential loss of a membrane during the course of evolution.
The process of secondary endosymbiosis is not unique to chlorarachniophytes. Excavates and chromalveolates also have secondary plastids. In the Excavates, secondary endosymbiosis of green algae led to euglenid protists. Meanwhile, in the Chromalveolates, secondary endosymbiosis of red algae led to the evolution of plastids in dinoflagellates, apicomplexans, and stramenopiles.
Case Study: What Does “Slug Power” Have to Do with the Origins of Chloroplasts?

Read the following excerpt from the Howard Hughes Medical Institute (HHMI) BioInteractive module entitled “Slug Power.”
The sacoglossan sea slug Elysia crispata can be found sunbathing on Caribbean reefs (Figure 20). The slug feeds on green algae but can survive for more than a month without eating. This is because sea slugs store chloroplasts, organelles in the cells of plants and algae that capture energy from sunlight and convert it to chemical energy by photosynthesis, as they ingest different species of green algae. The chloroplasts are stored in the slug’s digestive epithelium and remain active for up to 3–4 months, providing nutrients from photosynthesis, as well as camouflage by making the slug green in colour. “Kleptoplasty,” or “stolen plastids,” is the term for the slugs’ remarkable ability. Some marine protists including foraminifera, dinoflagellates, and ciliates are capable of kleptoplasty, but sea slugs are the only animals to exhibit kleptoplasty.
Think about it! Propose a hypothesis to explain how the ability of sea slugs to generate food from stolen chloroplasts relates to the evolution of the organelle itself.
Self-Check
Case Study Conclusion: What does “slug power” have to do with the origins of chloroplasts?
Show/Hide answer
They represent a living, powerful model system for studying the evolution of photosynthesis in eukaryotes through multiple endosymbiotic events. Since sea slugs can absorb chloroplasts for their own use, maybe something caused the ancestor of the eukaryotic cell to absorb bacteria capable of photosynthesis and gain their abilities. This supports the origins of chloroplasts from free-living prokaryotes and illustrates a physical arrangement that benefits both parties.
Learning Activity: Endosymbiotic Theory
Answer the following questions:
- Describe the hypothesized steps in the origin of eukaryotic cells.
Show/Hide answer
Eukaryotic cells arose through endosymbiotic events that gave rise to the energy-producing organelles within the eukaryotic cells, such as mitochondria and chloroplasts. The nuclear genome of eukaryotes relates the most closely to the Archaea, so it may have been an early archaean that engulfed a bacterial cell (alpha-proteobacterium) that evolved into a mitochondrion. Mitochondria appear to have originated from an alpha-proteobacterium, whereas chloroplasts originated as a cyanobacterium. There is also evidence of secondary endosymbiotic events. Other cell components may also have resulted from endosymbiotic events.
- Draw a flow chart describing the symbiotic adventure that led to the establishment of mitochondria (i.e., the endosymbiotic theory).
Show/Hide answer
Archean host cell + aerobic bacterium → atmospheric changes cause the aerobic bacterium to be engulfed by the host cell → the host cell provided a home for the bacterium without competition, and the bacterium used nutrients and oxygen to produce energy → over time, the relationship became required → aerobic bacterium lost genes needed to function apart from the host cell, resulting in the mitochondria.
- The amount of protein composing a plasma membrane, in comparison to a phospholipid, differs by organism. Bacteria have a very high protein-to-phospholipid ratio, around 2.5:1, while eukaryotes exhibit a ratio of 1:1, at least in their cell membrane. But remember that eukaryotes have multiple plasma membranes, one for every organelle. The protein-to-phospholipid ratio for their mitochondrial membrane is 2.5:1. What does this suggest about the relationship between mitochondria and bacteria?
Show/Hide answer
A similar protein/phospholipid ratio between mitochondria and bacterial plasma membranes contributes to the evidence that eukaryotes evolved from a bacterial ancestor.
✮ Learning Activity: Endosymbiotic Theory Online Interactive
The following online interactive will further familiarize you with the Endosymbiotic Theory. The emphasis is on the ‘process of science’ and how scientific evidence is evaluated. The interactive also highlights the importance of diversity within the scientific community of researchers. You were first introduced to this aspect while studying the history of the Cell Theory.
✮ Just a note that this activity may be one you choose to contribute as part of the Unit 1 assignment. For more details on this assignment, go to your Moodle Course Shell.
- Go to the article “Endosymbiosis: Cells within cells” and read through all 15 pages. Take the ‘side trips’ to explore the scientific method and more. Take note of the ‘side trip’ “Why diversity within the scientific community is so important in overcoming biases.”
- Answer the following questions:
- State two hypotheses for the origin of mitochondria and chloroplasts. State a prediction for each of these two hypotheses.
- There were multiple pieces of evidence supporting the endosymbiotic theory. List two strong pieces of evidence that helped lead to the ‘smoking gun’ in support of the endosymbiotic theory. How does this show that science builds?
- Given all your previous readings, would you fully agree with the following statement as evidence of the independence of mitochondria and chloroplasts? Please briefly justify your answer.
“They have their own ribosomes and so can make their own proteins.”
Key Concepts and Summary
- Cells come in many different shapes. Cells with different functions often have different shapes.
- Although cells come in diverse shapes, all cells have certain parts in common. These parts include the plasma membrane, cytoplasm, ribosomes, and DNA.
- Cells are classified as prokaryotic or eukaryotic based on their degree of internal compartmentalization.
- The prokarya (Eubacteria) are a diverse group of organisms, occupying almost every wet, dry, hot or cold nook and cranny of our planet. Despite this diversity, all prokaryotic cells share many structural and functional metabolic properties with each other and with eukaryotes.
- All prokaryotes have plasma membranes, cytoplasm, ribosomes, a cell wall, DNA, and lack membrane-bound organelles. Many also have polysaccharide capsules. Prokaryotic cells range in diameter from 0.1–5.0 µm.
- Prokaryotes lack an organized nucleus and other membrane-bound organelles. Prokaryotic DNA is found in a central part of the cell called the nucleoid.
- Like a prokaryotic cell, a eukaryotic cell has a plasma membrane, cytoplasm, and ribosomes, but a eukaryotic cell is typically larger than a prokaryotic cell. Eukaryotic cells tend to be 10 to 100 times the size of prokaryotic cells.
- Eukaryotic cells are defined by having a nucleus containing the DNA genome and bound by a nuclear membrane (or nuclear envelope) composed of two lipid bilayers that regulate the transport of materials into and out of the nucleus through nuclear pores.
- Eukaryotic cell morphologies vary greatly and may be maintained by various structures, including the cytoskeleton, the cell membrane, and/or the cell wall.
- As seen with ribosomes, shared structural and functional properties support the common ancestry of all life.
- Endosymbiotic theory states that mitochondria and chloroplasts (organelles found in many types of organisms) originate from bacteria. Significant structural and genetic information supports this theory.
- Endosymbiosis is the concept of one cell engulfing another and both cells benefiting from the relationship.
- Endosymbiosis was originally considered after observing the similarity between plant chloroplasts and free-living cyanobacteria.
- Although eukaryotic cells contain 80S ribosomes in the rough endoplasmic reticulum (membrane-bound ribosomes) and cytoplasm (free ribosomes), they contain 70s ribosomes in mitochondria and chloroplasts.
- Over time, endosymbionts may have transferred some of their DNA to the host nucleus, thus becoming dependent on the host for survival and completing full integration into a single organism.
- Finally, not only do eukaryotes share common ancestry with prokaryotes, but they also share living arrangements with them.
Long Descriptions
Figure 6 Image Description: The phylogenetic tree of life is divided into three branches: bacteria, archaea, and eukaryota. Bacteria include aquifex, green filamentous bacteria, planctomyces, cyanobacteria, proteobacteria, spirochetes, and gram positives. Archaea includes pyrodicticum, thermoproteus, methanococcus, methanosarcina, and halophiles. Eukaryota includes slime moulds, animals, fungi, plants, ciliates, entamoebae, microsporidia, and diplomonads. [Back to Figure 6]
Figure 7 Image Description: A prokaryotic cell is made up of three layers (from outside to inside): capsule, cell wall, and plasma membrane. Covering the outside of the capsule are fimbriae. Inside the plasma membrane is the cytoplasm. Cytoplasm contains different-sized dots (from smallest to largest): plasmid, ribosome, and inclusion. In the middle are squiggly lines representing a nucleoid. Extending out from the cytoplasm to outside the cell are two strings: a shorter one called pilus and a longer one called flagellum. [Back to Figure 7]
Figure 8 Image Description: A eukaryotic cell is made up of many organelles floating in the cytoplasm and contained within the plasma membrane. The plasma membrane is covered in thick lines called cilia. In the centre of the cell is a large nucleus made of nuclear pores, a nuclear envelope, and a nucleolus. Around the nucleus and taking up most of the cell are the connected lines of the rough endoplasmic reticulum covered in dots representing ribosomes. The other organelles scattered in the remaining space are smooth endoplasmic reticulum (rounded version of the rough endoplasmic reticulum), centrosomes (ridged cylinders), mitochondria (bean-shaped blobs), microfilaments (squiggly lines), microtubules (straight lines), lysosomes (small dots), peroxisomes (large dots), and a Golgi complex (ridges lined up close together). A long line string called a flagellum extends out of the cell from the cytoplasm. [Back to Figure 8]
Figure 11 Image Description: The ribosome’s large subunit contains tRNA with the dots of a growing peptide chain extending from it. A dotted line of mRNA separates the large subunit from the small subunit. [Back to Figure 11]
Figure 12 Image Description: In prokaryotes, ribosome large subunits are 50S and have 23S rRNA, 5S rRNA, and 31 ribosomal proteins. Small subunits are 30S and have 16S rRNA and 21 ribosomal proteins. In eukaryotes, ribosome large subunits are 60S and have 28S rRNA, 5S rRNA, 5.8S rRNA, and 50 ribosomal proteins. Small subunits are 40S and have 18S rRNA and 33 ribosomal proteins. [Back to Figure 12]
Figure 13 Image Description: A typical eukaryotic animal cell is made up of the following organelles:
- Nucleus, made up of:
- Nucleus envelope – membrane enclosing the nucleus. Protein-lined pores allow material to move in and out.
- Chromatin – DNA plus associated proteins.
- Nucleolus – a condensed region where ribosomes are formed.
- Cytoskeleton, made up of:
- Microtubules – form the mitotic spindle and maintain cell shape.
- Centrosome – microtubule-organizing centre.
- Intermediate filaments – fibrous proteins that hold organelles in place.
- Endoplasmic reticulum, classified as:
- Rough – associated with ribosomes; makes secretory and membrane proteins.
- Smooth – makes lipids.
- Vacuole.
- Mitochondira – produce energy.
- Cytoplasm.
- Golgi apparatus – modifies proteins.
- Lysosome – digests food and waste materials.
- Plasma membrane.
- Microfilaments – fibrous proteins; form the cellular cortex. [Back to Figure 13]
Figure 14 Image Description: A typical eukaryotic plant cell is made up of the following organelles:
-
- Plasmodesmata – channels connect two plant cells.
- Cell wall – maintains cell shape.
- Plasma membrane.
- Cytoplasm.
- Central vacuole – filled with cell sap that maintains pressure against the cell wall.
- Cytoskeleton – microtubules intermediate filaments microfilaments.
- Chloroplast – site of photosynthesis.
- Plastid – store pigments.
- Peroxisome.
- Mitochondria.
- Golgi apparatus.
- Ribosomes.
- Nucleus – contains chromatin, a nuclear envelope, and a nucleolus, as in an animal cell.
- Rough and smooth endoplasmic reticulum. [Back to Figure 14]
Figure 15 Image Description: The endosymbiotic theory includes three important events. First, the infoldings in the plasma membrane of an ancestral (proto-eukaryote) cell gave rise to endomembrane components, including a nucleus and endoplasmic reticulum. Second, in the first endosymbiotic event, the ancestral eukaryote consumed aerobic bacteria that evolved into mitochondria. Third, in a second endosymbiotic event, the early eukaryote consumed photosynthetic bacteria that evolved into chloroplasts. The second event results in a modern heterotrophic eukaryote, and the third event results in a modern photosynthetic eukaryote. [Back to Figure 15]
Media Attributions
- Figure 1: Figure 4.1 from OpenStax Biology 2e (Clark et al. 2018), is used under a CC BY Attribution license.
- Figure 2: Micrographia [uploaded by Alejandro Porto under “Hooke-Microscope-cork”] by Robert Hooke (1635-1702), via Wikimedia Commons, is in the public domain.
- Figure 3: Figure 3.6 from OpenStax Microbiology (Parker et al. 2016) is used under a CC BY 4.0 license.
- Figure 4: 2.2: Common Parts of the Cell is shared under a CK-12 license and was authored, remixed, and/or curated by CK-12 Foundation via edited source content.
- Figure 5: Basic-Components-of-a-cell by Christine Miller (2020) is used under a CC0 1.0 license.
- Figure 6: Phylogenetic tree [vector adaptation of Carl Woese at NASA Astrobiology Institute, 1990] by Eric Gaba (2006), via Wikimedia Commons, is in the public domain.
- Figure 7: Figure 2.10 from OpenStax Allied Health Microbiology (Bruslind 2019) is used under a CC BY-SA 4.0 license.
- Figure 8: Figure 3.35 from OpenStax Microbiology (Parker et al. 2016) is used under a CC BY 4.0 license.
- Figure 9: Figure 1.1.2.2 from BSC 3271: Microbiology for Health Sciences Sp21 (Kagle 2023) in LibreTexts Biology is used under a CC BY 4.0 license.
- Figure 10: Figure 3.36 from OpenStax Microbiology (Parker et al. 2016) is used under a CC BY 4.0 license.
- Figure 11: Figure 6 from BIS 2A: Introductory Biology (Singer) in LibreTexts Biology is used under a CC BY-SA 4.0 license.
- Figure 12: Difference in ribosomal ‘size’ illustration from Basic Cell and Molecular Biology (Bergtrom) in LibreTexts Biology is used under a CC BY-SA 4.0 license.
- Figure 13: Figure 4.8 from OpenStax Biology 2e (Clark et al. 2018) is used under a CC BY 4.0 license.
- Figure 14: Figure 4.8 from OpenStax Biology 2e (Clark et al. 2018) is used under a CC BY 4.0 license.
- Figure 15: Figure 3.7 from OpenStax Microbiology (Parker et al. 2016) is used under a CC BY 4.0 license.
- Figure 16: Figure 23.2 from OpenStax Biology 2e (Clark et al. 2018) is used under a CC BY 4.0 license.
- Figure 17: Figure 23.3 from OpenStax Biology 2e (Clark et al. 2018) is used under a CC BY 4.0 license.
- Figure 18: Figure 23.4 from OpenStax Biology 2e (Clark et al. 2018) is used under a CC BY 4.0 license.
- Figure 19: Figure 23.6 from OpenStax Biology 2e (Clark et al. 2018) is used under a CC BY 4.0 license.
- Figure 20: Elysia clarki, Florida by millerse (millerse 2014), via Wikimedia Commons, is used under a CC BY 3.0 license.
References
Antony van Leeuwenhoek (1632-1723). [date unknown]. Berkeley (CA): University of California Museum of Paleontology; [accessed 2024 Jan 29]. https://ucmp.berkeley.edu/history/leeuwenhoek.html.
Archaea. [date unknown]. Palo Alto (CA): CK-12 Foundation; [accessed 2023 Dec 9]. https://www.ck12.org/biology/archaea/.
Augenblick A, Thomas L. The wacky history of cell theory – Lauren Royal-Woods. TED-Ed. 2012 Jun 4, 6:11 minutes. [accessed 2024 Jan 16]. https://ed.ted.com/lessons/the-wacky-history-of-cell-theory.
Barnhill AE, Brewer MT, Carlson SA. 2012. Adverse effects of antimicrobials via predictable or idiosyncratic inhibition of host mitochondrial components. Antimicrob Agents Chemother. 56(8):4046–4051. https://journals.asm.org/doi/10.1128/aac.00678-12. doi:10.1128/aac.00678-12.
Berg OG, Kurland CG. 2000. Why mitochondrial genes are most often found in nuclei. Mol Biol Evol. 17(6):951–961. https://academic.oup.com/mbe/article/17/6/951/1037844. doi:10.1093/oxfordjournals.molbev.a026376.
Bergtrom G. 2021. Basic cell and molecular biology. Milwaukee (WI): University of Wisconson-Milwaukee; [accessed 2024 Jan 16]. https://bio.libretexts.org/Bookshelves/Cell_and_Molecular_Biology/Book%3A_Basic_Cell_and_Molecular_Biology_(Bergtrom).
Bergtrom G. 2021. Basic cell and molecular biology. Milwaukee (WI): University of Wisconson-Milwaukee; [accessed 2024 Jan 16]. Chapter 1.4: tour of the eukaryotic cell, Image 6 [difference between Eukaryotic and prokaryotic ribosomes]. https://bio.libretexts.org/Bookshelves/Cell_and_Molecular_Biology/Book%3A_Basic_Cell_and_Molecular_Biology_(Bergtrom)/01%3A_Cell_Tour_Lifes_Properties_and_Evolution_Studying_Cells/1.04%3A_Tour_of_the_Eukaryotic_Cell.
Biology Simulations. 2022. Cell structure [Interactive simulation]. Cambridge (MA): LabXchange; [accessed 2024 Jan 16]. https://www.labxchange.org/library/items/lb:LabXchange:6eb3b516:lx_simulation:1.
Bruslind L. 2019. General microbiology. OR: Oregon State University Ecampus; [accessed 2024 Jan 17]. https://open.oregonstate.education/generalmicrobiology/.
Bruslind L, OpenStax. 2019. Allied health microbiology. Corvallis (OR): Oregon State University Ecampus; [accessed 2024 Jan 16]. Figure 2.10. https://open.oregonstate.education/microbiology/chapter/3-3unique-characteristics-of-prokaryotic-cells/.
Clark MA, Choi J, Douglas M. 2018. Biology 2e. 2nd ed. Houston (TX): OpenStax. [accessed 2023 Sep 28]. https://openstax.org/books/biology-2e/pages/1-introduction.
Clark MA, Choi J, Douglas M. 2018. Biology 2e. 2nd ed. Houston (TX): OpenStax. [accessed 2023 Sep 28]. Figures 4.8, 23.2, 23.3. https://openstax.org/books/biology-2e/pages/1-introduction.
Corning Museum of Glass. Lens making in the 1600s [Video]. YouTube. 2016 Apr 28, 4:16 minutes. [accessed 2024 Jan 29]. https://www.youtube.com/watch?v=2SJY0foypAo.
Domain. [date unknown]. Palo Alto (CA): CK-12 Foundation; [accessed 2023 Dec 9]. https://www.ck12.org/biology/domain/.
Douglas AE. 2009. The microbial dimension in insect nutritional ecology. Funct Ecol. 23(1):38-47. https://besjournals.onlinelibrary.wiley.com/doi/10.1111/j.1365-2435.2008.01442.x. doi:10.1111/j.1365-2435.2008.01442.x.
Embley TM, Martin W. 2006. Eukaryotic evolution, changes and challenges. Nature. 440:623-630. https://www.nature.com/articles/nature04546. doi:10.1038/nature04546.
Fowler S, Roush R, Wise J. 2013. Concepts of biology. Houston (TX): OpenStax; [accessed 2024 Jan 17]. https://openstax.org/books/concepts-biology/pages/1-introduction.
Fowler S, Roush R, Wise J. 2013. Concepts of biology. Houston (TX): OpenStax; [accessed 2024 Jan 17]. Table 3.1. https://openstax.org/books/concepts-biology/pages/3-3-eukaryotic-cells.
Gaba E, Woese C. 2006. Phylogenetic tree [Image]. Wikimedia Commons; [updated 2023 Sep 24; accessed 2024 Jan 29]. https://commons.wikimedia.org/wiki/File:Phylogenetic_tree.svg.
Harris H. The birth of the cell. New Haven (CT): Yale University Press. p. 133.
Heins G. [date unknown]. A typical animal cell [Interactive simulation]. Madison (WI): Wisc-Online; [accessed 2024 Jan 29]. https://www.wisc-online.com/learn/natural-science/life-science/ap11403/a-typical-animal-cell.
Porto A, Hooke R. 2014. Hooke-Microscope-cork [Image]. Wikimedia Commons; [updated 2022 Dec 5; accessed 2024 Jan 29]. https://commons.wikimedia.org/wiki/File:Hooke-Microscope-cork.jpg.
Jaynes JM, Vernon LP. 1982. The cyanelle of Cyanophora paradoxa: almost a cyanobacterial chloroplast. Trends Biochem Sci. 7(1):22-24. https://www.sciencedirect.com/science/article/abs/pii/0968000482900585?via%3Dihub. doi:10.1016/0968-0004(82)90058-5.
Kagle J. 2023. BSC 3271: microbiology for health sciences sp21 (Kagle). Mansfield (PA): Mansfield University of Pennsylvania; [accessed 2024 Jan 16]. https://bio.libretexts.org/Courses/Mansfield_University_of_Pennsylvania/BSC_3271%3A_Microbiology_for_Health_Sciences_Sp21_(Kagle).
Kagle J. 2023. BSC 3271: microbiology for health sciences sp21 (Kagle). Mansfield (PA): Mansfield University of Pennsylvania; [accessed 2024 Jan 16]. Chapter 1.1.2: types of microorganisms. https://bio.libretexts.org/Courses/Mansfield_University_of_Pennsylvania/BSC_3271%3A_Microbiology_for_Health_Sciences_Sp21_(Kagle)/01%3A_Introduction/1.01%3A_An_Invisible_World/1.1.02%3A_Types_of_Microorganisms.
Khan Academy. Parts of a cell [Video]. YouTube. 2010 Feb 8, 21:03 minutes. [accessed 2024 Jan 16]. https://www.youtube.com/watch?v=Hmwvj9X4GNY.
Khan Academy. Cell theory | structure of a cell | biology | Khan Academy [Video]. YouTube. 2015 Jun 21, 8:02 minutes. [accessed 2024 Jan 16]. https://www.youtube.com/watch?v=zk3vlhz1b6k.
Kisch B. 1954. Forgotten leaders in modern medicine: Valentin, Gruby, Remak, Auerbach. Trans Am Philos Soc. 44(2):139-317. https://www.jstor.org/stable/1005758. doi:10.2307/1005758.
MeriSTEM. Leeuwenhoek’s microscope [Video]. LabXchange. 2021 Nov 8, 5:48 minutes. [updated 2023 Jul 3; accessed 2024 Jan 29]. https://www.labxchange.org/library/items/lb:LabXchange:2d4b0230:video:1.
Miller C. 2020. Human biology. Kamloops (BC): Thompson Rivers University; [accessed 2024 Jan 17]. https://humanbiology.pressbooks.tru.ca/. [Adapted from the CK-12 Foundation textbook College Human Biology by Jean Brainard and Rachel Henderson].
Miller C. 2020. Human biology. Kamloops (BC): Thompson Rivers University; [accessed 2024 Jan 17]. Figure 4.2.4. https://humanbiology.pressbooks.tru.ca/chapter/4-2-discovery-of-cells-and-cell-theory/.
millerse. 2014. Elysia clarki, Florida [Image]. Wikimedia Commons; [updated 2016 Mar 2, accessed 2026 May 14]. https://commons.wikimedia.org/wiki/File:Elysia_clarki,_Florida.jpg.
Parker N. Schneegurt M, Thi Tu A-H, Lister P, Foster BM. 2016. Microbiology. Houston (TX): OpenStax; [accessed 2024 Jan 17]. https://openstax.org/books/microbiology/pages/1-introduction.
Parker N. Schneegurt M, Thi Tu A-H, Lister P, Foster BM. 2016. Microbiology. Houston (TX): OpenStax; [accessed 2024 Jan 17]. Chapter 3.2: foundations of modern cell theory, Science and plagiarism. Figures 3.6, 3.7, 3.35, 3.36. https://openstax.org/books/microbiology/pages/1-introduction.
Retraction Watch. 2010. New York (NY): The Center for Scientific Integrity; [updated 2024 Jan 17; accessed 2024 Jan 17]. https://retractionwatch.com/.
Sagan L. 1967. On the origin of mitosing cells. J Theor Biol. 14(3):225-274. https://www.sciencedirect.com/science/article/abs/pii/0022519367900793?via%3Dihub. doi:10.1016/0022-5193(67)90079-3.
SERP Institute. 2021. Comparing four cells: animal, plant, bacteria, yeast [interactive simulation]. Cambridge (MA): LabXchange; [accessed 2014 Jan 16]. https://www.labxchange.org/library/items/lb:LabXchange:fbdb87f4:lx_simulation:1.
Singer M. 2019. BIS 2A: introductory biology (Singer). Davis (CA): University of California, Davis; [accessed 2024 Jan 16]. https://bio.libretexts.org/Courses/University_of_California_Davis/BIS_2A_(2018)%3A_Introductory_Biology_(Singer).
Singer M. 2019. BIS 2A: introductory biology (Singer). Davis (CA): University of California, Davis; [accessed 2024 Jan 16]. Master resources, Eukaryotic cell: structure and function. https://bio.libretexts.org/Courses/University_of_California_Davis/BIS_2A_(2018)%3A_Introductory_Biology_(Singer)/MASTER_RESOURCES/Eukaryotic_Cell%3A_Structure_and_Function*.
Stewart D [Username: The Doc]. 2014. Robert Hooke [blog]. Famous Scientists. [accessed 2023 Dec 8]. https://www.famousscientists.org/robert-hooke/.
Understanding Science Team. 2022. Endosymbiosis: cells within cells. Berkeley (CA): University of California Museum of Paleontology; [accessed 2024 Jan 16]. https://undsci.berkeley.edu/cells-within-cells-an-extraordinary-claim-with-extraordinary-evidence/.
Understanding Science Team. 2022. The scientific community: diversity makes the difference. Berkeley (CA): University of California Museum of Paleontology; [accessed 2024 Jan 16]. https://undsci.berkeley.edu/understanding-science-101/the-social-side-of-science-a-human-and-community-endeavor/the-scientific-community-diversity-makes-the-difference/.
Wakim S, Grewal M. 2022. Human biology (Wakim & Grewal). Oroville (CA): Butte College; [accessed 2024 Jan 17]. https://bio.libretexts.org/Bookshelves/Human_Biology/Human_Biology_(Wakim_and_Grewal).
Wakim S, Grewal M. 2022. Human biology (Wakim & Grewal). Oroville (CA): Butte College; [accessed 2024 Jan 16]. Table 5.3.3. https://bio.libretexts.org/Bookshelves/Human_Biology/Human_Biology_(Wakim_and_Grewal)/05%3A_Cells/5.03%3A_Variation_in_Cells.
Webster C. 1981. Biology medicine and society 1840-1940. Cambridge (England): Cambridge University Press. p. 118-119.
Wikipedia Contributors. 2023. Antonie van Leeuwenhoek. Wikipedia, The Free Encyclopedia; [updated 2023 Dec 8; accessed 2023 Dec 8]. https://en.wikipedia.org/w/index.php?title=Antonie_van_Leeuwenhoek&oldid=1188946567.
Wikipedia Contributors. 2023. Robert Hooke. Wikipedia, The Free Encyclopedia; [updated 2023 Dec 8; accessed 2023 Dec 8]. https://en.wikipedia.org/w/index.php?title=Robert_Hooke&oldid=1188835504.
Wikipedia Contributors. 2023. Theodor Schwann. Wikipedia, The Free Encyclopedia. [updated 2023 Oct 12; accessed 2023 Dec 8]. https://en.wikipedia.org/w/index.php?title=Theodor_Schwann&oldid=1179860086.
Zuchora-Walske C. 2015. Key discoveries in life sciences. Minneapolis (MN): Lerner Publishing Group. p. 12-13.

{kind=link}
{kind=link}